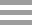

近日,Plant Biotechnology Journal杂志在线发表了由太阳成集团tyc33455cc李冰冰课题组撰写的“FvMAPK6-Mediated FvMYB44s/FvSWEET1 Dual-Layer Regulation Modulates Sugar Accumulation in Strawberry Fruit, With FvSPS3 Enabling Quality–Yield Balance” 论文。该研究通过整合分子生物学和遗传学方法,揭示了蛋白激酶FvMAPK6作为磷酸化枢纽,在草莓果实中应答蔗糖信号协同调控糖分与花青素积累的分子机制,阐明了磷酸化介导的果实糖积累双调控新通路,并筛选出可实现果实品质与产量平衡的关键靶基因FvSPS3,为培育高糖稳产的草莓新品种提供了核心基因资源与创新育种思路。

可溶性糖是决定果实品质的关键,既可作为初生代谢物影响风味与甜度,又能作为信号分子调控果实成熟过程(如色素积累)。目前,果实成熟中调控糖分积累与糖信号转导的分子机制仍不清晰,且高糖优质与高产之间的遗传负相关问题,已成为果实品质育种中的瓶颈问题。草莓果实中主要糖类为蔗糖、葡萄糖和果糖,其中蔗糖不仅是决定不同草莓果实品种可溶性固形物含量的关键,还可作为信号同时促进糖分与花青素积累,但其介导的信号组分与通路尚不明确。李冰冰课题组的这项研究,解析了FvMAPK6-FvMYB44/FvMAPK6-FvSWEET1 双通路应答蔗糖信号、协同调控草莓果实品质形成的分子机制,丰富了果实糖信号转导与成熟调控的理论体系,为解决果实品质育种中“糖-产量”拮抗的经典难题,提供了全新解决方案。全文主要研究结果如下:
1. FvMAPK6正调控草莓果实中糖和花青素的协同积累
研究团队以二倍体草莓(F. vesca cv. Ruegen)为材料,创制了FvMAPK6稳定过表达转基因株系(FvMAPK6-OE)和CRISPR/Cas9介导的基因编辑转基因株系(Fvmapk6-cr)。结合果实的成熟进程观察和生理生化指标检测发现,与野生型(WT)相比,FvMAPK6-OE果实的花青素积累显著加速,葡萄糖、果糖、蔗糖三类可溶性糖的积累模式与花青素一致(图1A、1C-F);而Fvmapk6-cr果实的花青素与可溶性糖积累均显著延迟(图1B、1D-F)。以上结果表明,FvMAPK6在协同调控草莓果实糖和花青素的积累中发挥关键作用。
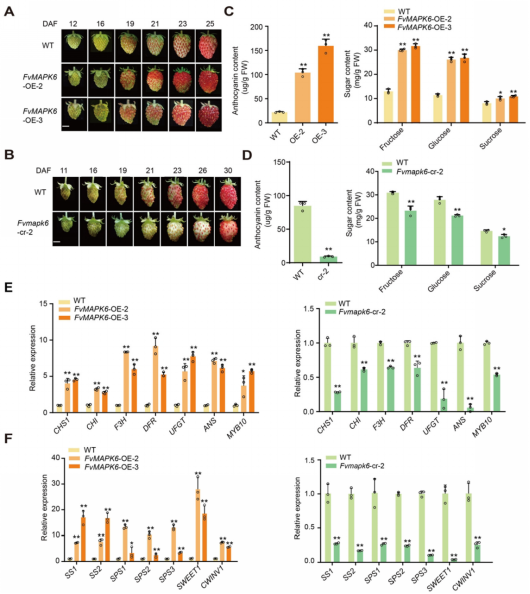
图1 FvMAPK6调控草莓果实中糖和花青素的协同积累
2. FvMAPK6通过磷酸化促进FvMYB44s降解并减弱其转录活性
为进一步阐明FvMAPK6介导FvMYB44磷酸化的生物学意义,作者研究了FvMAPK6 对FvMYB44.1和FvMYB44.2蛋白水平及转录活性的影响。半体外蛋白降解试验表明,FvMAPK6可显著促进FvMYB44.1和FvMYB44.2通过蛋白酶体途径降解(图2A);内源蛋白检测发现,FvMYB44.1在果实成熟过程中蛋白水平持续下降(图2B),且在FvMAPK6-OE果实中显著降低,在Fvmapk6-cr果实中显著升高,与蛋白降解结果一致(图2C)。进一步通过草莓果实内源转录报告试验发现,FvMAPKK4–FvMAPK6磷酸化级联可分别通过磷酸化FvMYB44.1和FvMYB44.2的Ser62和Ser226位点,部分解除其对下游靶基因的转录抑制作用(图 3D‑F)。该结果阐明了FvMAPK6在转录水平调控糖积累的分子途径。
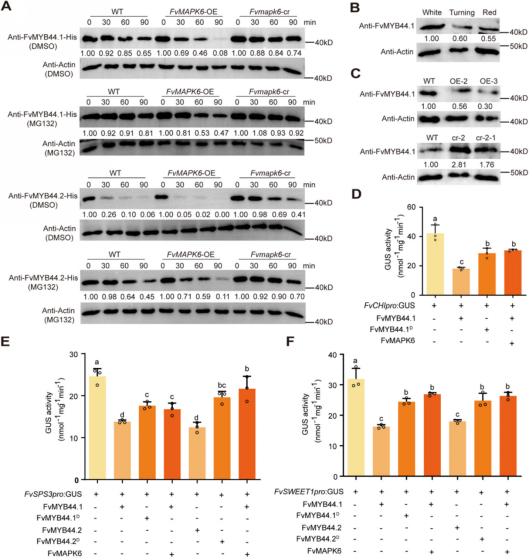
图2 FvMAPK6促进转录因子FvMYB44.1/FvMYB44.2的蛋白降解,并解除二者对下游基因的抑制作用
3. FvMAPK6增加FvSWEET1的蛋白丰度,并通过磷酸化修饰改变其转运活性
除了通过磷酸化转录因子来调控下游结构基因的表达外,蛋白激酶还可直接磷酸化功能蛋白以发挥其生物学功能。作者对FvMAPK6转基因果实进行了磷酸化蛋白质组学分析,发现,FvSWEET1的磷酸化水平发生显著变化。且FvSWEET1是SWEET家族中表达量最高的成员,其表达也受FvMAPK6显著调控。通过制备FvSWEET1抗体,进行蛋白水平检测显示:与野生型相比,FvMAPK6显著促进FvSWEET1蛋白积累(图3A),这与FvMAPK6通过调控FvMYB44.1解除其对下游靶基因FvSWEET1的抑制结果一致。己糖/糖转运缺陷型酵母菌株中异源表达FvSWEET1及烟草原生质体定位结果表明,FvSWEET1为质膜己糖转运蛋白(图3B、3C)。此外,在烟草叶片和草莓果实中均检测到FvMAPK6与FvSWEET1的相互作用(图3D-F)。随后作者探究了FvMAPK6对FvSWEET1磷酸化介导的调控机制。Phos‑tag试验显示,FvSWEET1的磷酸化水平受到FvMAPK6显著调控(图3G)。将FvSWEET1 C端第223位丝氨酸(Ser‑223)磷酸化位点分别突变为天冬氨酸(D,模拟磷酸化形式)和丙氨酸(A,非磷酸化形式)。与FvSWEET1相比,FvSWEET1D突变体的葡萄糖转运活性增强,而FvSWEET1A突变体转运活性减弱(图3H、3I)。以上结果表明,FvMAPK6介导的磷酸化对维持FvSWEET1的转运活性至关重要。
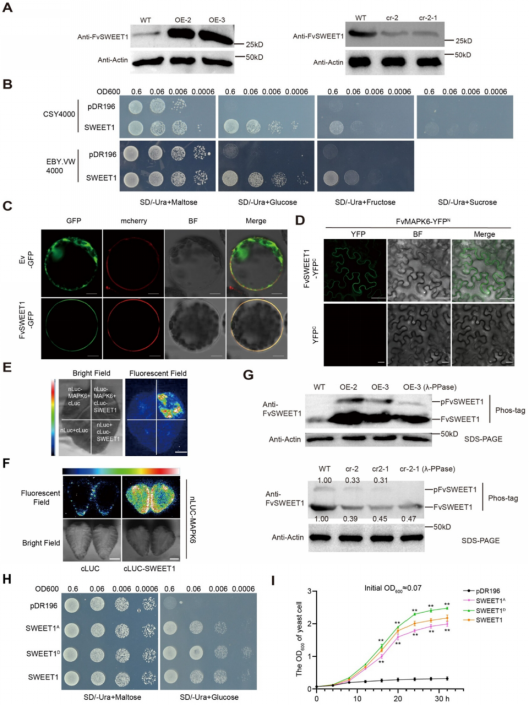
图3 FvMAPK6磷酸化己糖转运蛋白FvSWEET1以调控其功能
4. FvSPS3可精准提升草莓果实中的糖含量,且不影响植株生长与产量
超表达FvMAPK6虽能显著提高草莓果实含糖量,但会导致植株矮化、产量下降;相反,Fvmapk6-cr双等位突变体植株的生长势与产量均高于野生型(图4A、4B)。上述结果表明,FvMAPK6通过多效性作用调控生长/产量与果实品质之间的权衡关系,直接调控其表达无法实现品质与产量的协同提升。为规避这一权衡矛盾,作者在FvMAPK6调控网络中,筛选出果实中高表达、且与蔗糖积累直接相关的下游特异靶基因FvSPS3。创制FvSPS3超表达株系后发现,该株系植株形态与结果性状均与野生型无显著差异(图4C–E),但其果实中花青素与糖分积累均显著加快(图4F、4G);进一步在商品成熟期(花青素含量一致)果实中检测发现,FvSPS3‑OE果实的含糖量显著高于野生型(图4H、4I)。综上,FvSPS3 是可实现草莓果实品质提升与产量稳定的特异性靶基因,为精准育种提供了核心靶点。
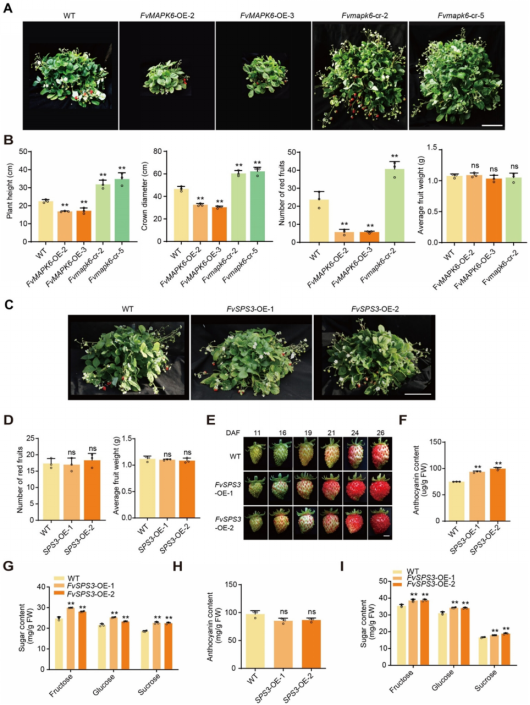
图4 FvMAPK6调控草莓的营养生长和产量,FvSPS3调控草莓果实的花青素和糖分含量且不影响植株的生长和产量
综上所述,在草莓果实发育过程中,FvMAPK6表现出高水平的磷酸化,蔗糖处理进一步增强其磷酸化水平。具体而言,蔗糖激活的FvMAPK6与FvMKK4形成磷酸化级联反应,直接磷酸化转录抑制因子FvMYB44.1和FvMYB44.2,通过促进二者蛋白降解并降低二者的转录活性来削弱其抑制功能,从而解除对糖分积累和花青素生物合成的抑制。此外,FvMAPK6增加己糖转运蛋白FvSWEET1的蛋白丰度及其磷酸化介导的转运活性,进一步增强糖分积累。总之,FvMAPK6作为一种蔗糖响应性激酶,通过双重机制协调糖分和花青素积累:(1)由FvMYB44.1/44.2直接磷酸化介导的转录途径;(2)由FvSWEET1磷酸化介导的翻译后途径。实线箭头表示本研究中建立的调控关系;虚线箭头表示基于现有和先前证据的假定机制。FvMAPK6产生的内源性蔗糖作为信号分子直接放大下游生物学事件的潜在机制有待未来研究阐明(图5)。此外,由于对信号调控网络上游组分进行操纵往往会产生多效性影响,易造成植株生长与产量下降。该研究通过精准筛选,确定了下游关键功能基因FvSPS3。结果表明,对FvSPS3进行靶向调控,能够在不影响植株正常生长与产量的前提下,有效提高果实糖分积累,为优质高产育种提供了更为安全、高效的分子靶点。
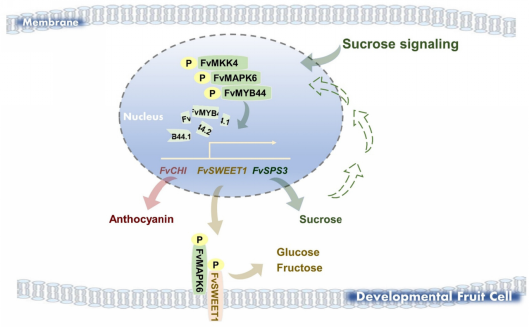
图5 蔗糖激活的FvMAPK6调控草莓果实中糖分与花青素积累的模型
太阳成集团tyc33455博士生冯倩倩为该论文第一作者,太阳成集团tyc33455李冰冰教授为通讯作者。研究工作得到了北京林业大学孟冬教授、浙江农林大学范臻教授、江苏省农业科学院袁华招副研究员的支持和帮助。研究工作得到了国家重点研发计划、国家自然科学基金等项目的资助。
供图、供稿:李冰冰课题组
编辑:张月夕 蒋晓彤
审核:付国强